نوع مطالعه: تحقیقی |
موضوع مقاله: طب رزم دریافت: 1403/6/17 | ویرایش نهایی: 1403/12/22 | پذیرش: 1403/8/6 | انتشار: 1403/10/1
متن کامل فارسی: (1388 مشاهده)
مقدمه
فرایند طبیعی پیری با از دست دادن تدریجی هموستاز همراه است که به تغییرات فیزیولوژیک متنوعی در عملکرد سلولها و بافتها منجر میشود [1]. با افزایش سن و سالمندی استرس اکسیداتیو افزایش مییابد، این موضوع به اختلال در عملکرد عضلات و آتروفی عضلانی منجر میشود. برای کنترل استرس اکسایشی سلولها با توقف چرخه سلولی از طریق فعال کردن نشانگرهای پیری، پروتئینهای سرکوبگر تومور[1] p53 و p16 از آسیب سلولی جلوگیری میکنند [2]. پروتئینهای p53 و p16 بهعنوان نشانگرهای پیری شناختهشدهاند و مطالعات نشان داده که همزمان با افزایش سن بیان این پروتئینها افزایش مییابد. p53 فرایندهای زیستی مانند پاسخ استرس، چرخه سلولی، تکثیر، پیری و آپوپتوز را تنظیم میکند. عملکردهای متنوع p53 در تنظیم سوختوساز بافت چربی در مطالعات مورد توجه قرار گرفته است که p53 را با ناهنجاریهای سوخت و سازی مشاهدهشده در پیری، چاقی، التهاب و سرطان مرتبط میکند [3]. p53 همچنین موجب القای پروتئینهای درگیر در مسیر آپوپتوز، پروتئینهای شبکه آندوپلاسمی، کاسپاز-6 و کاسپاز -9 میشود. علاوه بر این p53 موجب سرکوب ژن ضدآپوپتوزی مانند Bcl-2 میشود. بنابراین p53 در سطح رونویسی از طریق افزایش بیان ژنهای آپوپتوزی و کاهش بیان ژنهای ضدآپوپتوزی موجب تحریک آپوپتوز در سلولهای توموری میشود. پژوهشها نشان دادهاند چاقی ناشی از رژیم غذایی پرچرب و بینظمی در سوختوساز چربی در بسیاری از سلولها مانند بافت چربی، آئورت، پانکراس و کبد سبب افزایش بیان ژن p53، p16، p19 و p21 میشود [4]. بیان ژن p16 در بیشتر سلولهای پیر، مهارکننده کیناز وابسته به سیکلین و سرکوبکننده تومور صورت میگیرد که سبب توقف رشد میشود. گزارششده است که p16 ممکن است نقشی در تنظیم رونویسی فاکتورهای وابسته به پیری در سلولها داشته باشد [5]. فعالیت ورزشی یک مداخله در سبک زندگی با اثرات ضدپیری شناخته شده است که قادر به خنثی کردن چندین نشانه پیری از جمله التهاب مرتبط با افزایش سن است. در مدل موشهای پیر، فعالیت ورزشی با کاهش فعالیت GTT[2]و سطوح p53، p21 و IL-6، نشانگرهای پیری را سرکوب و واسطههای التهابی را کاهش میدهد [6]. فعالیتهای ورزشی با تقویت دفاع اکسایشی در بدن منجر به کاهش التهاب سیستمی در موشهای چاق میشود. ورنر[3] و همکاران گزارش کردند که فرایند پیری تحت تأثیر تمرینات ورزشی از طریق کاهش سطوح p53 وp16 آئورت موشها، قرار میگیرد [7]. گزارش شده است که فعالیت ورزشی حاد موجب کاهش p53 هستهای به طور مستقیم و یا از طریق تنظیم مثبت Nrf2 که منجر به غیرفعال شدن مسیرهای سیگنالینگ p53-P21Cip1 و p16INK4a-RB میشود. با توجه به این اثرات متنوع در شرایط آزمایشگاهی، توضیح نقش عملکردی در داخل بدن p53 در طول پیری دشوار بوده است [8]. این احتمال وجود دارد که پاسخ تا حدی به محل سلولی آن و همچنین مدت و شدت محرک بستگی داشته باشد. ارتباط بین آپوپتوز و فعالیت در موشهای دیابتی در پژوهش قربانزاده و همکاران بررسی شد و کاهش معنیدار p53 گزارش شد [9]. سیدقمی و همکاران تغییر معنیدار p53 در عضله اسکلتی پس از تمرینات استقامتی مشاهده نکردند. به نظر میرسد شدت و نوع تمرینات میتواند تأثیرگذار باشد. تمرین تناوبی با شدت بالا (HIIT)[4]باعث افزایش کاردیومیوپاتی دیابتی از طریق سرکوب آپوپتوز کاردیومیوسیت با واسطه miR-1 میشود [10]. عبداللهی دیبا و همکاران گزارش کردند که تمرینات استقامتی تأثیر معنیداری بر بیان ژنهای سیتوکروم C وp53 در عضله قلب موشهای صحرایی نر نداشت [11]همچنین رشواناسماعیل و همکاران گزارش کردند که اثر 12 هفته تمرین هوازی بر فاکتورهای بیان ژنهای p53 و miR-34a در آزمودنیهای سالم معنیدار نبود [12]. درک تأثیر HIIT بر این عوامل در افراد مسن چاق برای توسعه مداخلات ورزشی هدفمند برای بهبود نتایج سلامتی در این جمعیت مهم است. درحالی که برخی شواهد حاکی از یک اثر سنولیتیک[5] (قادر به کشتن سلولهای پیر) بالقوه HIIT بر روی p53 و p16 است، مکانیسمهای خاص و میزان این اثر در افراد مسن چاق نامشخص است [13]. HIIT به دلیل اثرات بالقوه آن بر عوامل مرتبط با افزایش سن در افراد چاق مورد توجه قرارگرفته است. چندین مطالعه نشان دادهاند که HIIT ممکن است اثر ضدپیری داشته باشد و بهطور بالقوه بیان p53 و p16 را کاهش دهد. نظامدوست و همکاران گزارش کردند که بیان p53 پس از چهار هفته تمرین تناوبی شدید افزایش مییابد [14]. با این حال، اثرات خاص HIIT بر p53 و p16 در افراد مسن چاق بهطور کامل مشخص نشده است [10،15] از طرفی دیگر اختلالات انسولینی و بیماری دیابت در جمعیت سالخورده دارای شیوع بالایی است. از آنجایی که امروزه بیماریهای مزمن از قبیل انواع دیابت و عوارض ناشی از آن رو به افزایش است، مقاومت به انسولین ناشی از مصرف کالری بیش از حد، فعالیت ورزشی ناکافی و کمبود ترشح انسولین از سلولهای β ممکن است در ایجاد دیابت نوع دو تأثیرگذار باشند. بنابراین پانکراس برای رفع نیاز بدن شروع به تکثیر سلولهای β کرده که نتیجه آن سالمندی این سلولها است [16]. بیماریهای متابولیکی مانند دیابت نوع دو بهعنوان نتیجهای از استعداد ژنتیکی و عوامل محیطی است که شامل رژیم غذایی پرکالری، چاقی و سبک زندگی بیتحرک هستند. با توجه به نقش p53 با ایجاد مقاومت به انسولین از طریق تأثیر بر مسیرهای متابولیکی مانند متابولیسم گلوکز و در نتیجه القای ژنهای مسیر سیگنالی گیرندههای انسولین، انجام پژوهشهایی برای بررسی تأثیر رژیم غذایی پرچرب، که منتهی به چاقی میشود و تأثیر فعالیت ورزشی بر این عوامل ضروری به نظر میرسد. پس با توجه به موارد مطرح شده این امر ضروری است که به بررسی دقیق تأثیر HIITبر عملکرد پانکراس و جلوگیری از بیماریهای التهاب مزمن پانکراس و سرطان پانکراس پرداخت. یافتههای این مطالعه ممکن است بینش ارزشمندی در مورد مزایای بالقوه HIIT برای کاهش پیری سلولی مرتبط با سن و آسیبشناسیهای مرتبط با آن در جمعیت مسن چاق ارائه دهد. در پژوهشهای گذشته تأثیر روشهای مختلف تمرینی بر موارد ذکر شده بررسی و نتایج متفاوتی گزارششده است، همچنین با بررسی متون علمی موجود مشاهده گردید که مطالعات محدودی درباره بیان p53 و p16 در بافت پانکراس انجامشده است. بنابراین هدف از مطالعه حاضر بررسی تأثیر هشت هفته تمرینات HIIT بر بیان ژن p53 و p16 در بافت پانکراس موشهای تغذیه شده با غذای پرچرب بود.
روش بررسی
در این مطالعه تجربی که با طرح پسآزمون همراه با گروه کنترل انجام شد، 20 سر موش صحرایی نر نژاد ویستار 52 هفتهای در قفسهای استاندارد در دمای 2±22 درجه سانتیگراد نگهداری شدند. چرخه تاریکی (12 ساعت روشنایی و 12 ساعت تاریکی) بهطور دقیق رعایت شد. بعد از گذشت یک هفته از زمان سازگاری با محیط آزمایشگاه، حیوانات بر اساس همسانسازی وزنی به چهار گروه تقسیم شدند: گروه غذای نرمال، گروه غذای نرمال + تمرین، گروه غذای پرچرب و گروه غذای پرچرب + تمرین. حیوانات به آب و غذا بهصورت آزاد دسترسی داشتند و با یکی از دو رژیم غذایی پرچرب (60% چربی) یا رژیم غذایی استاندارد تغذیه شدند. توان هوازی به این صورت برآورد شد که در ابتدا رتها به مدت پنج دقیقه بر روی تردمیل با سرعت شش متر بر دقیقه و شیب صفر درجه گرم کردند. سپس هر سه دقیقه، سرعت تردمیل بهطور تدریجی 3 متر بر دقیقه افزایش مییافت تا زمانی که حیوانات به واماندگی و خستگی میرسیدند و قادر به ادامه تمرین نبودند. ملاک اصلی برای رسیدن به VO2max، عدم توانایی رتها در ادامه دادن پروتکل تمرینی با افزایش سرعت بود. بهعبارتدیگر، زمانی که رتها نتوانستند تمرین را ادامه دهند و به واماندگی رسیدند، میزان VO2max آنها را تخمین زده شد [17]. برای آشنایی و سازگاری با HIIT رتها به مدت 2 هفته بر روی تردمیل تمرین داده شدند، در ابتدای هفته اول از سازگاری با تمرین رتها با سرعت 10 متر بر دقیقه و شیب صفر درجه به مدت 10 دقیقه شروع به تمرین کردند و در آخر هفته دوم با همان سرعت 10 متر بر دقیقه، مدتزمان تمرین به 30 دقیقه رسید. رتها به دو گروه کلی HIIT و گروه کنترل تقسیم شدند که با توجه به طرح پژوهش، گروه تمرین به مدت 8 هفته تحت تمرین قرار گرفتند. برنامه پروتکل HIIT، توجه به پژوهشهای قبلی [18] بدینصورت طراحی شد که 3 روز در هفته و هر روز یک جلسه 22 دقیقهای HIITشامل 4 دقیقه دویدن با شدت 45 تا 55% VO2max، 9 تناوب 1 دقیقهای با شدت 90 تا 95% VO2max و 1 دقیقه دویدن بین هر تناوب با شدت 50% VO2max بود. 48 ساعت پس از آخرین جلسه تمرینی و بعد از 12 ساعت ناشتایی موشهای صحرایی با تزریق درون صفاقی ترکیبی از کتامین (50 میلیگرم بر کیلوگرم) و زایلازین (5 میلیگرم بر کیلوگرم) بیهوش شدند سپس با خارج کردن قلب قربانی شدند و به سرعت بافت پانکراس جداسازی شد و بعد از شستشو با سرم فیزیولوژی و جداکردن قسمتهای زاید، به نیتروژن مایع انتقال یافته و سپس در دمای منفی 80 درجه سانتیگراد تا زمان انجام آزمایش نگهداری شد. جهت بررسی بیان ژنها با استفاده از Real-time PCR شرکت ماکروژن[6] (سئول، کره جنوبی) تمام پرایمرها توسط نرمافزار Allele IDv7.8 طراحی شد و از ژن β2m (بتا 2 میکروگلوبولین) بهعنوان کنترل داخلی استفاده گردید. تمام پرایمرها به صورتاتصال اگزون-اگزون طراحی شدند (جدول 1). جهت اطمینان از عدم تکثیر DNA ژنومی از 25 نانوگرم cDNA و 25 نانوگرم RNA در تیوبهای جداگانه از واکنش PCR و به کارگیری از ژل آگاروز 1.5% استفاده شد. تکثیر cDNA و مشاهده باند مورد انتظار توسط پرایمر اختصاصی و عدم تکثیر RNA پس از واکنش PCR نمایانگر عدم تکثیر DNA ژنومی است. سپس برای هر یک از پرایمرها کارایی PCR اندازهگیری و منحنی استاندارد برای آنها رسم گردید.
ملاحظات اخلاقی
در مطالعه حاضر، استفاده از حیوانات آزمایشگاهی منطبق بر دستورالعملهای بینالمللی مراقبت و استفاده از حیوانات آزمایشگاهی بود. کلیه مراقبتها و اصول اخلاقی بهطور کامل بر اساس دستورالعملهای استفاده و مراقبت از حیوانات آزمایشگاهی رعایت شدند. پژوهش حاضر با تأیید کمیته اخلاق اجرا گردید.
تجزیه و تحلیل آماری
در این تحقیق دادهها با استفاده از میانگین و انحراف معیار ارائه شدهاند. همچنین برای بررسی نحوه توزیع دادهها از آزمون شاپیرو-ویلک استفاده شد. در ادامه با توجه به پارامتریک بودن دادهها برای بررسی تفاوت بین گروهی از آزمون تحلیل واریانس یکراهه استفاده شد. همچنین برای تعیین محل تفاوت بین گروهها از آزمون تعقیبی بونفرونی استفاده شد. دادههای تحقیق حاضر در نرمافزار SPSS نسخه 26 تجزیهوتحلیل شدند. همچنین سطح معنیداری برای تمام محاسبات 0.05 در نظر گرفته شد.
یافتهها
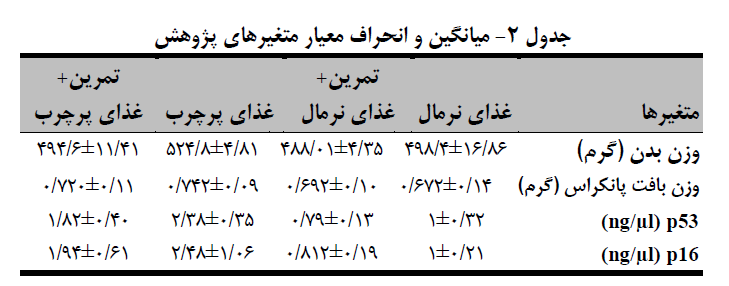
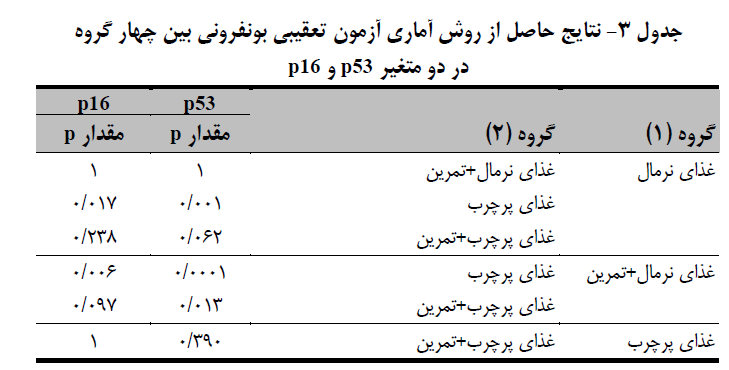
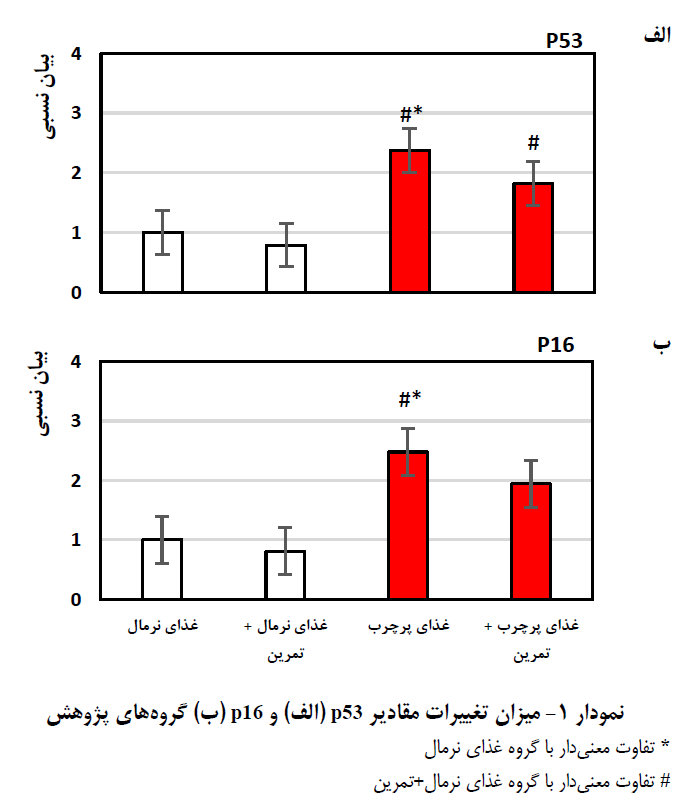
میانگین و انحراف معیار متغیرهای پژوهش در جدول 2 آمده است. یافتههای مربوط به بیان ژن حاکی از این بود که مقادیر بیان ژن p53 بین چهار گروه مختلف تفاوت معنیداری وجود دارد (0.001=p)، این تفاوت بین گروه غذای نرمال با گروه غذای پرچرب (0.001=p)، گروه غذای نرمال+تمرین با غذای پرچرب+تمرین (0.013=p) و گروه غذای نرمال+ تمرین با گروه غذای پرچرب (0.0001=p) تفاوت معنیدار وجود دارد (جدول 3 و نمودار 1-الف). میزان معنیداری آزمون تحلیل واریانس یکطرفه (0.003p=) نشان داد که ازلحاظ فاکتور p16 نیز بین چهار گروه مختلف تفاوت معنیداری وجود دارد. نتایج آزمون تعقیبی نشان داد که مقادیر بیان ژن p16 بین گروه غذای نرمال با گروه غذای پرچرب (0.017=p) و گروه غذای نرمال+ تمرین با گروه غذای پرچرب (0.006=p) تفاوت معنیدار وجود دارد (جدول 3 و نمودار 1-ب).
بحث و نتیجهگیری
نتایج مطالعه حاضر نشان داد که تمرین تناوبی شدید منجر به کاهش معنادار در بیان نسبی ژنهای p16 و p53 در بافت پانکراس موشهای سالمند تغذیه شده با غذای پرچرب میشود؛ درصورتیکه در گروههای کنترل (بدون تمرین با غذای نرمال و غذای پرچرب)، میزان بیان این ژنها در سطح بالاتری بود. پروتئین p53 بهعنوان یک عامل مولکولی کلیدی پیشنهاد شده است که سوبستراهای متابولیسم و بیوژنز میتوکندری ناشی از تمرین در عضلات اسکلتی را تنظیم میکند. اختلال در محتوا و عملکرد میتوکندری با آسیبهای زیادی مانند اختلالات متابولیک، افزایش سن، دیابت نوع 2، چاقی و سرطان و همچنین کاهش عملکرد تمرینی همراه است. همسو با نتایج پژوهش حاضر، دشتیان و همکاران گزارش کردند که تمرینات تداومی و تناوبی شدید، بیان ژنهای سرکوبگر توموری p53 و PTEN را کاهش میدهد و مصرف مکمل ویتامین E به همراه این نوع تمرینات میتواند تا حدودی باعث اثرات متفاوت در بیان این ژنها شود [15]. در پژوهش دیگری جان بزرگی و همکاران تأثیر هشت هفته تمرین هوازی متعاقب 5 هفته القای چاقی با رژیم غذایی پرچرب بر بیان پروتئینهای سالمندی p16 و p53 بافت پانکراس موشهای دیابتی را بررسی کردند. نتایج این مطالعه حاکی از کاهش معنیداری در بیان ژنهای p16 و p53 متعاقب تمرین هوازی با شدت متوسط بود. بر اساس یافتههای این پژوهش تمرین هوازی احتمالاً با کاهش عوامل مربوط به سالمندی سلولی نظیر p16 و p53 در سلولهای بتا موجب بهتر شدن حساسیت انسولینی و به تأخیر افتادن سالمندی سلولی ناشی از دیابت میشود [16]. در مقابل عبداللهی دیبا و همکاران گزارش کردند که تمرینات استقامتی تأثیر معنیداری بر بیان ژنهای سیتوکروم C وp53 در عضله قلب موشهای صحرایی نر نداشت [11]. همچنین رشواناسماعیل و همکاران گزارش کردند که اثر 12 هفته تمرین هوازی بر فاکتورهای بیان ژنهای p53 و miR-34a در آزمودنیهای سالم معنی دار نبود [12]. این که چگونه ورزش یا تمرین، بیان این ژن را در درازمدت کاهش میدهد مورد بحث است. گزارش شده است که تمرین منظم، استرس اکسیداتیو را کاهش میدهد و دفاع آنتیاکسیدانی بدن را تقویت میکند [19]. بنابراین با توجه به اینکه یکی از دلایل افزایش بیان p53 افزایش استرس اکسیداتیو است، میتوان انتظار داشت که سطح بیان ژن p53 به دلیل کاهش استرس اکسیداتیو کاهش یابد [15]. تأثیر مثبت فعالیتهای ورزشی بر پارامترهای متعددی از سلامت جسمی، قلبی و متابولیک در موشهای میانسال و مهمتر از آن، غلبه بر اثرات مخرب اضافی مواد مغذی نشان داده شده است. در پیری و چاقی، بافت چربی منبع اصلی واسطههای التهابی است که در پیدایش دیابت و سایر بیماریهای مزمن نقش دارد [20]. شفر و همکاران در پژوهشی القای p16 و p53 را در بافت چربی احشایی و القای p21 را در چربی احشایی و زیرجلدی و کبد در موشهای 12 ماهه که در 16 یا 30 هفته قبل مداخلهای با رژیم غذایی سرشار از قند و چربی و/یا ورزش دریافت کردند، مشاهده کردند. به نظر میرسد بیان این نشانگرها در سایر بافتها از جمله پانکراس نیز افزایش مییابد [21]. آنها بیان کردند که نتایج ترکیبی به اتفاق آرا نشان میدهد که مواد غذایی اضافی منجر به القای پیری در بافتهای متعددی میشود که مسئول هماهنگی سلامت متابولیک هستند. در پژوهش حاضر نیز افزایش نشانگرهای پیری در گروه رژیم غذایی پرچرب در مقایسه با غذای نرمال مشاهده شد که میتواند تأثیرات رژیم پرچرب بر القای پیری در بافت پانکراس را نشان دهد. گزارش شده است که تغذیه با چربی بالا، بیان p16INK4a مرتبط با افزایش سن را که توسط فراوانی mRNA در کبد یا طحال تعیین میشود، تسریع میکند. علاوه بر این، مصرف بیش از حد مواد غذایی ممکن است تجمع سلولهای پیر p16INK4a و/یا p53 مثبت را در بافت چربی القا کند [21]. پیری سلولی با توقف رشد برگشتناپذیر مشخص میشود. افزایش بیان دو تنظیمکننده کلیدی چرخه سلولی مهاری، p16Ink4a و p21cip1 و سرکوبگر تومور p53، نشانگرهای اصلی فنوتیپ ترشحی مرتبط با پیری (SASP)[7]در نظر گرفته میشوند [22]. SASP واسطه التهاب مزمن با درجه پایین است که پیری بیولوژیکی و شروع بیماریهای مرتبط با سن را تسریع میکند [23]. سالمندان چاق تقویت SASP را از طریق تنظیم مثبت در فعالیت p53، p16 و p21 نشان میدهند که خطر ابتلا به اختلالات قلبی عروقی، عصبی و متابولیک را افزایش میدهد [24]. HIIT از طریق افزایش آنتیاکسیدانها و سطح آنزیم ترمیم DNA، واکنش آسیب DNA را سرکوب میکند و تجمع هستهای p53 را کاهش میدهد [25]. همین سازگاریها همچنین ممکن است از طریق مهار رونویسی مشابه، با تنظیم مثبت p16 و p21 مخالفت کنند. گزارش شده است که p53 در تنظیم مسیرهای سیگنالینگ وابسته به میتوکندری اضافی، از جمله اتوفاژی/میتوفاژی و آپوپتوز نقش دارد. نقش p53 در تنظیم آپوپتوز شناختهشده است، زیرا میتواند ژنهای پروآپوپتوز متعددی از جمله Bax و Bid را به صورت رونویسی تنظیم کند تا آسیب DNA را القا کند [3]. علاوه بر این، p53 خود میتواند در سطح میتوکندری قرار گیرد، جایی که میتواند سینتیک منافذ انتقال نفوذپذیری را تنظیم کند [26]. قبلاً نشان داده شده بود که فعالیت ورزشی مزمن باعث کاهش نسبت Bax:Bcl-2 همزمان با کاهش سیتوکروم c و آزادسازی پروتئین AIF23 میشود که نشاندهنده سازگاریهای ضد آپوپتوز در میتوکندری است [27]. یوسف و همکاران تأثیر HIIT و تمرین مداوم با شدت متوسط به مدت 12 هفته بر بیان ژن بافت چربی، ترکیب بدن و نشانگرهای زیستی خون در سالمندان چاق بررسی کردند. نتایج این مطالعه نشان میدهد که HIIT استراتژیهای مداخلهای مؤثر در افراد مسن چاق هستند، به نظر میرسد HIIT اثرات مفیدتری دارد. به طور خاص، HIIT منجر به پیشرفتهای بالاتری نسبت به MICT در کاهش وزن، ظرفیتهای عملکردی، توده بدون چربی و نشانگرهای عضله اسکلتی محتوای میتوکندری، همجوشی و میتوفاژی شد [4]. علاوه بر این، HIIT آزادسازی IL-6 را تحریک میکند [28]، که بیانp16 را با سرکوب فعالیت سیگنالینگ p38 MAPK/NF kB کاهش میدهد [29]. تأثیر HIIT بر این مسیرها ممکن است زمینهساز کاهش مشاهدهشده در التهاب ناشی از SASP و نرخ پیری بیولوژیکی باشد. HIIT ترشح سیتوکین های التهابی را تسهیل میکند که نشان داده شده است سلولهای ایمنی را تحریک میکند و این امر به نوبه خود سلولهای پیر را برای آپوپتوز هدف قرار میدهند [30]. این تفکیک سلولهای آسیبدیده پایدار همچنین به کاهش التهاب مزمن زمینهای و توسعه عصبی کمک میکند. این یافتهها نشان میدهد که تمرین تناوبی شدید تأثیر مثبتی بر بیان فاکتورهای p53 و p16 در بافت پانکراس رتهای سالمند تغذیهشده با غذای پرچرب دارد، به نحوی که در گروههای که تمرین انجام دادند شاهد کاهش بیان فاکتورهای p53 و p16 در بافت پانکراس رتهای سالمند بودیم و در گروههای که غذای پرچرب مصرف کردند شاهد افزایش بیان فاکتورهای p53 و p16 در بافت پانکراس رتهای سالمند بودیم. این نتایج ارتقاء سلامت بافت پانکراس و کاهش اثرات منفی غذای پرچرب را نشان دهد.
تشکر و قدردانی
این پژوهش باکد اخلاق IR.ILAM.REC.1402.020 به تأیید کمیته اخلاق در پژوهش دانشگاه ایلام رسیده است. نویسندگان بر خود لازم می دانند که از همه افرادی که در طی انجام این تحقیق کمک کردند تشکر نمایند.
همه نویسندگان در ایدهپردازی و انجام طرح، همچنین نگارش اولیه مقاله یا بازنگری آن سهیم بودهاند و همه با تأیید نهایی مقاله حاضر مسئولیت دقت و صحت مطالب مندرج در آن را میپذیرند.
منابع مالی
در این پژوهش از هیچ ارگانی کمک مالی دریافت نگردید.